|
|
|
|
|
|||||||||||
|
Electronic Flora of South Australia Species Fact Sheet
Phylum Rhodophyta – Class Florideophyceae – Order Corallinales – Family Corallinaceae – Subfamily Lithophylloideae
Selected citations: Woelkerling & Campbell 1992: 41, figs 22–32.
Synonyms
Melobesia corallinae Crouan & Crouan 1867: 150, 252, pl. 20 gen 133 bis figs 6–11
Dermatolithon corallinae (Crouan & Crouan) Foslie in Børgesen 1902: 402.
Tenarea corallinae (Crouan & Crouan) Notoya 1974: 47 (invalid).
Titanoderma corallinae (Crouan & Crouan) Woelkerling et al. 1985: 333. Chamberlain 1991: 66, figs 3, 16–18, 52j, 208–224. Chamberlain & Irvine 1994b: 90, figs 34, 35.
Additional references involving these and other binomials are given by Woelkerling & Campbell 1992.
Thallus normally pinkish, encrusting to layered, warty, lumpy or fruticose, mostly 10–50 mm across and 0.2–1 (–5) mm thick or tall, epigenous and partially or completely affixed by cell adhesion; protuberant branches simple or branched, mostly 1–7 mm in diameter and 3–7 mm long; lamellate branches applanate and inconspicuous, mostly 3–12 mm across and 1–2 mm long. Structure pseudoparenchymatous, organisation dorsiventral in crustose portions and lamellate branches but radial in protuberant branches; construction dimerous, dimerous and monomerous or largely monomerous; dimerous portions consisting of a single ventral layer of branched, laterally cohering, filaments each composed of palisade and/or non-palisade cells 5–20 µm long and (13–) 20–65 (–82) µm high, and additionally of simple or branched, laterally cohering, filaments that arise dorsally and more or less perpendicularly from cells of ventral layer filaments and are each usually composed of cells 5–16 µm in diameter and 5–40 (–55) µm long; monomerous portions consisting of a single system of branched, laterally cohering, filaments that collectively contribute to a ventrally or centrally situated core and a peripheral region where portions of core filaments or their derivatives curve outwards towards the thallus surface, with each filament composed of cells 5–14 µm in diameter and 5–35 (–55) µm long; epithallial cells 4–9 µm in diameter and 2–6 [n long, terminating most filaments at the thallus surface, with distal walls rounded or flattened but not flared; cells of adjacent filaments joined by secondary pit-connections; cell-fusions and haustoria unknown; trichocytes rare, single.
Reproduction: Vegetative reproduction unknown. Gametangia, carposporangia, tetrasporangia and bisporangia produced in uniporate conceptacles; gametangia and carposporangia formed on thalli separate to those bearing tetrasporangia and bisporangia.
Gametangial thalli monoecious or dioecious. Carpogonia terminating 2- or 3-celled filaments arising from the female conceptacle chamber floor. Mature female-carposporangial conceptacle roofs flush with or somewhat protruding above surrounding thallus surface, 50–115 µm thick, composed of 5–12 layers of cells above the chamber, conceptacle chambers 185–233 (–305) µm in diameter and 60–110 (–140) µm high. Carposporophytes composed of a conspicuous central fusion cell and several-celled gonimoblast filaments bearing terminal carposporangia 27–40 (–57) µm in diameter. Spermatangial filaments unbranched, borne in groups of 1–3 on initials across the male conceptacle chamber floor, mature male conceptacle roofs flush with or slightly protruding above surrounding thallus surface, 25–40 µm thick, composed of 2–5 layers of cells above the chamber, male conceptacle chambers 115–145 µm in diameter and 30–50 µm high.
Tetrasporangial/bisporangial conceptacle roofs non- or slightly protruding above surrounding surface, mostly 2–4 cells thick above chamber, pore canals lined with somewhat projecting cells that do not completely occlude the canal, conceptacle chambers (155–) 190–235 (–280) µm in diameter and 68–105 (–136) µm high, floor of mature chambers usually 6 or more cells below thallus surface; tetrasporangia and bisporangia scattered across the conceptacle chamber floor or peripheral to a central columella, each mature sporangium (15–) 25–55 (–95) µm in diameter and (35–) 50–95 (–125) µm long, containing zonately arranged tetraspores or bispores.
Type from Brest, France; lectotype in CO (unnumbered); designated by Chamberlain (1991, p. 67); depicted in Chamberlain (1991, p. 64, fig. 208) and Woelkerling & Campbell (1992, p. 43, fig. 22A).
Selected specimens: Eyre ("Nine Mile Reef'), W. Aust., 2–3 m deep (Woelkerling, Platt & Jones, 1.ii.1984; LTB, 13974). Point Sinclair, S. Aust., 0–2 m deep (Woelkerling, Platt & Jones, 15.ii.1984; LTB, 14507). Elliston, S. Aust., 5 m deep (Turner, 28.x.1981; LTB, 15360). Vivonne Bay, Kangaroo I., S. Aust., low eulittoral (Womersley, 14.i.1947; AD, Al2978 = LTB, 13587 and Campbell & Penrose, 8.iv.1988; LTB, 15884). Penneshaw, Kangaroo I., S. Aust., 0–1.5 m deep (Campbell & Penrose, 13.iv.1988; LTB, 15669). Nora Creina, S. Aust. (Woelkerling, 6.xi.1987; LTB, 15578). Bridgewater Bay, Vic., lower eulittoral (Womersley, 21.viii.1953; AD, A19053 = LTB, 13646 and 14.iv.1959; AD, A22633 = LTB, 13621). Apollo Bay (Haylings Reef), Vic. (Pope & Bennett, 17.i.1949; AD, A15243 = LTB, 13703). Anglesea (Potato Patch Reef), Vic., 8–9 m deep (Beanland, 31.viii.1982; LTB, 12721). Eddystone Point, Tas., 0–3 m deep (Platt & Woelkerling, 23.ii.1983; LTB, 13171, 13172, 13174, 13176). Eaglehawk Neck (Tessellated Pavement), Tas., 1–2 m deep (Platt & Woelkerling, 26.ii.1983; LTB, 13236, 13240). Eaglehawk Neck (Clydes I.), Tas., 0–6 m deep (Platt, 26.ii.1983; LTB, 13306).
Distribution: Probably widespread; many records outside Europe, however, require confirmation [see Chamberlain (1991) and Woelkerling & Campbell (1992)].
In Australia, Eyre, W. Aust., to Anglesea, Vic., and the E coast of Tasmania.
Taxonomic notes: Lithophyllum corallinae has been found in southern Australia on rock, snails, limpets, abalone shells and sea urchins in intertidal pools, on reef edges and subtidally to depths of 9 m. Gametangial, carposporangial, tetrasporangial and bisporangial thalli can occur in the same population. Bisporangia and tetrasporangia, however, were not found on the same individual. All southern Australian thalli examined were dioecious and epiphytic populations were not found. In the British Isles, in contrast, both monoecious and dioecious thalli occur and only epiphytic collections are known (Chamberlain & Irvine 1994b, p. 93). Woelkerling & Campbell (1992, p. 97) report that, rarely, intermediates can occur between L. corallinae and L. frondosum (as L. bermudense) and conclude that further studies of the relationships of these two species are required.
References:
BØRGESEN, F. (1902). The marine algae of the Faeroes. In Botany of the Faeroes Part 2, pp. 339–532. (H.H. Tiele: Copenhagen.) Note: Also reprinted without change in pagination by Det. Nordiske Forlag, Copenhagen.
CHAMBERLAIN, Y.M. & IRVINE, L.M. (1994b). Lithophylloideae Setchell. In Irvine, L. M. & Chamberlain, Y. M. (Eds), Seaweeds of the British Isles. Volume 1 Rhodophyta Part 2B Corallinales, Hildenbrandiales pp. 58–112. (HMSO: London.)
CHAMBERLAIN, Y.M. (1991). Historical and taxonomic studies in the genus Titanoderma (Rhodophyta, Corallinales) in the British Isles. Bull. Br. Mus. Nat. Hist. (Bot.) 21, 1–80.
CROUAN, P.L. & CROUAN, H.M. (1867). Florule du Finistère. (F. Klincksieck: Paris.)
HEYDRICH, F. (1897b). Melobesieae. Ber. dt. bot. Ges. 15: 403–420, Plate 18.
NOTOYA, M. (1974). Spore germination in crustose coralline Tenarea corallinae, T. dispar and T. tumidula. Bull. Jap. Soc. Phycol. 22, 47–51.
WOELKERLING, W.J. & CAMPBELL, S.J. (1992). An account of southern Australian species of Lithophyllum (Corallinaceae, Rhodophyta). Bull. Br. Mus. Nat. Hist., Bot. Ser. 22, 1–107.
WOELKERLING, W.J., CHAMBERLAIN, Y.M. & SILVA, P.C. (1985). A taxonomic and nomenclatural reassessment of Tenarea, Titanoderma and Dermatolithon (Corallinaceae, Rhodophyta) based on studies of type and other critical specimens. Phycologia 24, 317–337.
The Marine Benthic Flora of Southern Australia Part IIIB complete list of references.
Publication:
Womersley, H.B.S. (28 June, 1996)
The Marine Benthic Flora of Southern Australia
Rhodophyta. Part IIIB. Gracilarialse, Rhodymeniales, Corallinales and Bonnemaisoniales
Reproduced with permission from The Marine Benthic Flora of Southern Australia Part IIIB 1996, by H.B.S. Womersley. Australian Biological Resources Study, Canberra. Copyright Commonwealth of Australia.
Illustrations in Womersley Part IIIA, 1996: FIGS 62E, 63G, 102, 103.
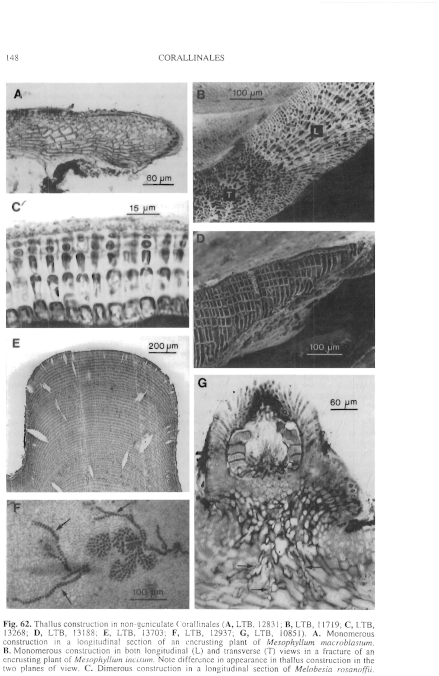
Figure 62 enlarge
Fig. 62. Thallus construction in non-geniculate Corallinales (A, LTB, 12831; B, LTB, 11719; C, LTB, 13268; D, LTB, 13188; E, LTB, 13703; F, LTB, 12937; G, LTB, 10851). A. Monomerous construction in a longitudinal section of an encrusting plant of Mesophyllum macroblastum. B. Monomerous construction in both longitudinal (L) and transverse (T) views in a fracture of an encrusting plant of Mesophyllum incisum. Note difference in appearance in thallus construction in the two planes of view. C. Dimerous construction in a longitudinal section of Melobesia rosanoffii.
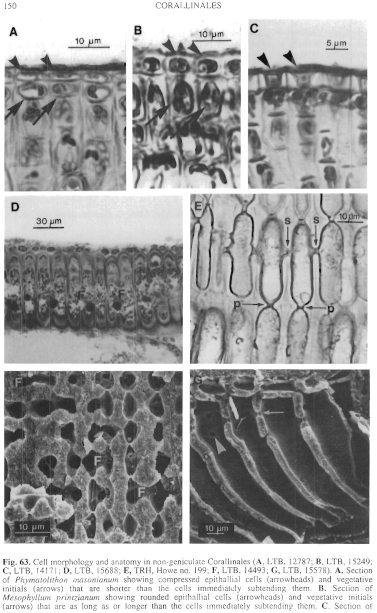
Figure 63 enlarge
Fig. 63. Cell morphology and anatomy in non-geniculate Corallinales (A, LTB, 12787; B, LTB, 15249; C, LTB, 14171; D, LTB, 15688; E, TRH, Howe no. 199; F, LTB, 14493; G, LTB, 15578). A. Section of Phymatolithon masonianum showing compressed epithallial cells (arrowheads) and vegetative initials (arrows) that are shorter than the cells immediately subtending them. B. Section of Mesophyllum printzianum showing rounded epithallial cells (arrowheads) and vegetative initials (arrows) that are as long as or longer than the cells immediately subtending them. C. Section of Sporolithon durum showing epithallial cells (arrow heads) with flared outer walls D. Palisade cells of contiguous filaments of Metamastophora flabellata showing cell-fusions (F). E. Columnar cells of Lithophyllum frondosum showing primary (arrow - p) and secondary (arrow - s) pit connections. F. Fracture of Sporolithon durum showing fusions (F) between cells of contiguous filaments. G. Fracture of Lithophyllum corallinae showing pit connections in surface (arrowhead) and sectional (arrow) views.
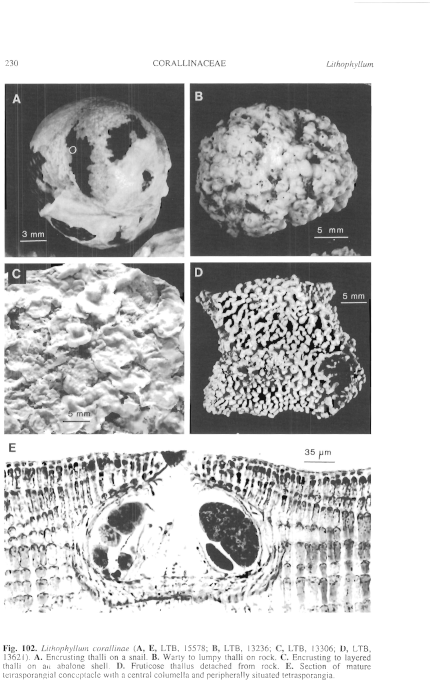
Figure 102 enlarge
Fig. 102. Lithophyllum corallinne (A, E, LTB, 15578; B, LTB, 13236; C, LTB, 13306; D, LTB, 13621). A. Encrusting thalli on a snail. B. Warty to lumpy thalli on rock. C. Encrusting to layered thalli on an abalone shell. D. Fruticose thallus detached from rock. E. Section of mature tetrasporangial conceptacle with a central columella and peripherally situated tetrasporangia.
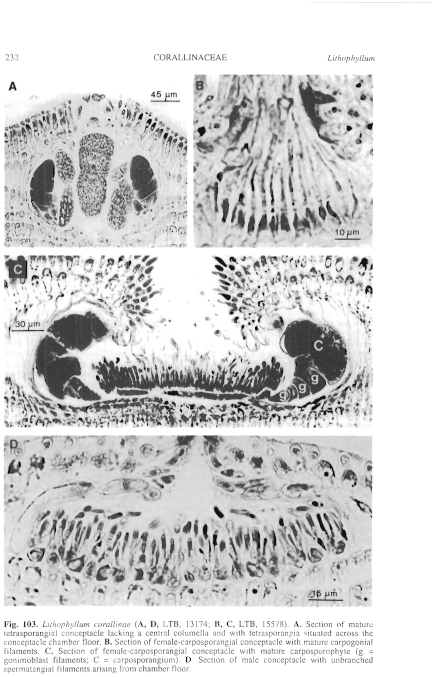
Figure 103 enlarge
Fig. 103. Lithophyllum corallinae (A, D, LTB, 13174; B, C, LTB, 15578). A. Section of mature tetrasporangial conceptacle lacking a central columella and with tetrasporangia situated across the conceptacle chamber floor. B. Section of female-carposporangial conceptacle with mature carpogonial filaments. C. Section of female-carposporangial conceptacle with mature carposporophyte (g = gonimoblast filaments; C = carposporangium). D. Section of male conceptacle with unbranched spermatangial filaments arising from chamber floor.

|
Email Contact: State Herbarium of South Australia |

|









