|
|
|
|
|
|||||||||||
|
Electronic Flora of South Australia Genus Fact Sheet
Phylum Phaeophyta – Order Chordariales – Family Chordariaceae
Thallus moderately to much branched, cylindrical, very mucoid, usually 10–30 cm long and 3–5 µm in diameter. Medulla multiaxial, of more or less parallel longitudinal filaments producing fairly abundant hyphae but not obscuring the longitudinal filaments, often becoming hollow. Subcortex extensive, often broader than the cortex, of branched, non-pigmented filaments arising at right angles from peripheral medullary filaments. Cortex of determinate assimilatory filaments arising in groups from the outer subcortical cells, filaments simple, often curved above, of short cells. Phaeophycean hairs present, arising from outer subcortical cells.
Thallus (Fig. 33C,D) grey-brown to dark brown, very mucoid, usually moderately branched with long laterals from near the base, to profusely branched with third or fourth order laterals and when young with short laterals near the apices, usually 10–30 cm long and (1–) 2–4 (–5) mm in diameter, cylindrical, becoming hollow, with a small discoid holdfast 1–3 mm across, epilithic. Basal system of juvenile plants consisting of a prostrate loose weft of irregularly globose, pigmented cells, from which erect medullary filaments develop. Medulla (Fig. 34F) of more or less parallel longitudinal filaments of cylindrical to slightly swollen cells 15–40 (–80) µm in diameter and L/B (1.5–) 2–8 (–12), from which numerous hyphae develop but without obscuring the longitudinal filaments; older parts often becoming hollow. Subcortex (Fig. 34E,F) extensive, often 200–700 µm broad and as wide or wider than the cortex, of branched, radiating filaments 10–18 µm in diameter near the medulla and 6–12 µm in diameter outwardly, cells L/B (2–) 4–10, developing from the peripheral medullary cells or hyphae, and with inner cells in turn producing hyphae. Cortical filaments (Fig. 34E,F) simple, determinate, borne in groups from upper subcortical cells or continuing as an inner branch of the subcortex, usually curved above, 100–200 (–250) µ and 12–20 cells long with upper cells ovoid (often swollen on upper side), 8–12 (–14) µm in diameter and L/B 1–2, with occasional intercalary divisions, lower cells cylindrical, 5–8 (–10) mm in diameter and L/B 2–3 (–5). Phaeoplasts (Fig. 34G) several per cell, discoid, each with a pyrenoid; cells with numerous small physodes. Phaeophycean hairs (Fig. 34F) frequent, arising from outer subcortical cells, 6–10 µm in diameter.
Reproduction: Reproduction by clavate to subspherical unilocular sporangia borne on outer subcortical cells at base of cortical filaments. Gametophytes filamentous, branched, dioecious, isogamous (in T. crassa — Peters & Müller 1986b) with the sporophyte also reproducing directly from plurilocular sporangia (Yotsui 1978) or from unilocular sporangia (Yotsui 1982).
Unilocular sporangia (Fig. 34F) clavate to ovoid to subspherical, sessile on outer subcortical cells and lying within the cortex, (40–) 50–90 µm long and (20–) 30–70 µm in diameter. Plurilocular sporangia unknown.
Life history diplohaplontic and heteromorphic (Yotsui 1978), or direct (Yotsui 1982).
Type: Type species: T. crassa (Suringar) Kylin.
Type from Georgetown, Tas., in Herb. Harvey, TCD.
Selected specimens: Sceale Bay, S. Aust., lower eulittoral (Bailey, 25.viii.1967; ADU, A31833). Victor Harbor, S. Aust., uppermost sublittoral just E. of causeway (Bailey, 6.xii.1980; ADU, A51934). Yilki, Victor Harbor, S. Aust., lower eulittoral pools near shore (Womersley, 7.ix.1985; ADU, A56720--Marine Algae of southern Australia" No. 256). Robe, S. Aust., low eulittoral (Womersley, 8.xii.1973; ADU, A44590). Apollo Bay, Vic., lower eulittoral pools (Bailey, 23.i.1967; ADU, A31754). Phillip I., Vic., lower eulittoral at Penguin Parade Beach (Sinkora A2237, 19.x.1975; ADU, A48506). Safety Cove, Port Arthur, Tas., lower eulittoral (Womersley, 31.x.1982; ADU, A55949).
Distribution: From Sceale Bay, S. Aust. to Port Phillip, Vic. and around Tasmania, usually on sand covered lower eulittoral rock during summer (August to late January).
Taxonomic notes: Tinocladia is distinguished by the extensive subcortex, radiating from the medulla of longitudinal filaments and bearing groups of cortical assimilatory filaments. The five species included by Kylin (1940) are not well separated morphologically and structurally, and may be only forms of T. australis (see also Mower & Widdowson 1969, pp. 74,75).
Tinocladia australis (Harvey) Kylin 1940: 34, pl. 6 fig. 14. Bailey & Womersley in Womersley 1967: 235.
Liebmannia australis Harvey I 859b: 291.
Eudesme harvevana Kylin: 31, pl. 5 fig. 13. Bailey & Womersley in Womersley 1967: 233.
Tinocladia australis is distinguished by the very wide subcortex, with some cortical filaments originating from mid cells of the subcortex. Eudesme harvevana is indistinguishable structurally from T. australis, and is interpreted as being younger plants with more abundant, short, upper laterals (Fig. 33D). This form intergrades into typical T. australis in the Victor Harbor region of South Australia.
References:
KYLIN, H. (1940). Die Phaeophyceenordnung Chordariales. Acta Univ. lund. N.F. Avd. 2, 36(9), 1–67, Plates 1–8.
MOWER, A. & WIDDOWSON, T.B. (1969). New records of marine algae from southern California. Bull. South. Calif Acad. Sci. 68, 72–81.
PETERS, A.F. & MÜLLER, D.G. (1986b). Critical re-examination of sexual reproduction in Tinocladia crassa, Nemacystis decipiens and Sphaerotrichia divaricata (Phaeophyceae, Chordariales). Jap. J. Phycol. 34, 69–73.
WOMERSLEY, H.B.S. (1967). A critical survey of the marine algae of southern Australia. II. Phaeophyta. Aust. J. Bot. 15, 189–270.
YOTSUI, T. (1978). On the life cycle of an edible brown alga, Tinocladia crassa. Bull. Jap. Soc. Sci. Fish. 44, 861–867.
YOTSUI, T. (1982). The life cycle of Tinocladia crassa (Suringar) Kylin (Phaeophyta, Chordariales) without a haploid gametophyte from Kuchinotsu, Kyushu, Japan. Jap. J. Phycol. 30, 113–115.
The Marine Benthic Flora of Southern Australia Part II complete list of references.
Publication:
Womersley, H.B.S. (14 December, 1987)
The Marine Benthic Flora of Southern Australia
Part II
©Board of the Botanic Gardens and State Herbarium, Government of South Australia
Illustrations in Womersley Part II, 1997: FIGS 33C,D, 34 E–G.
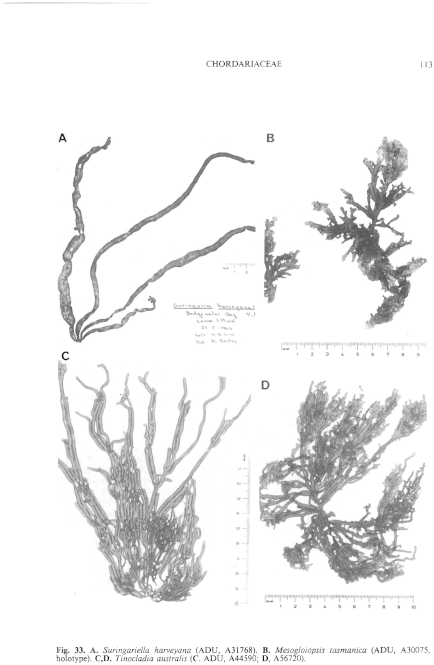
Figure 33 enlarge
Fig. 33. A. Suringariella harveyana (ADU, A31768). B. Mesogloiopsis tasmanica (ADU, A30075, holotype). C,D. Tinocladia australis ( C, ADU, A44590; D, A56720).
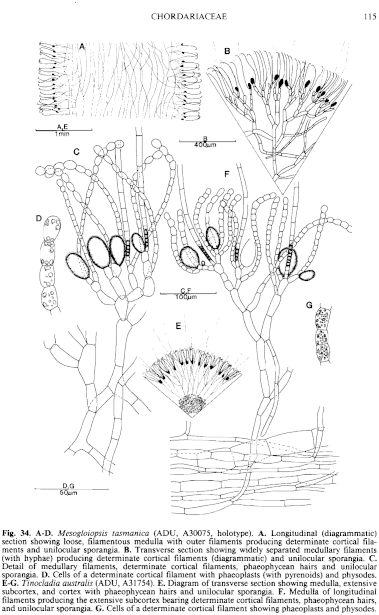
Figure 34 enlarge
Fig. 34. A–D. Mesogloiopsis tasmanica (ADU, A30075, holotype). A. Longitudinal (diagrammatic) section showing loose, filamentous medulla with outer filaments producing determinate cortical filaments and unilocular sporangia. B. Transverse section showing widely separated medullary filaments (with hyphae) producing determinate cortical filaments (diagrammatic) and unilocular sporangia. C. Detail of medullary filaments, determinate cortical filaments, phaeophycean hairs and unilocular sporangia. D. Cells of a determinate cortical filament with phaeoplasts (with pyrenoids) and physodes. E–G. Tinocladia australis (ADU, A31754). E. Diagram of transverse section showing medulla, extensive subcortex, and cortex with phaeophycean hairs and unilocular sporangia. F. Medulla of longitudinal filaments producing the extensive subcortex bearing determinate cortical filaments, phaeophycean hairs, and unilocular sporangia. G. Cells of a determinate cortical filament showing phaeoplasts and physodes.

|
Email Contact: State Herbarium of South Australia |

|









