|
|
|
|
|
|||||||||||
|
Electronic Flora of South Australia Species Fact Sheet
Phylum Phaeophyta – Order Chordariales – Family Splachnidiaceae
Selected citations: Asensi & Delépine 1976: 35, figs 1–3. Harvey 1858: pl. 14. Kuckuck 1929: 77, figs 110–114. Mitchell & Whitting 1892: 1, pls 1–3. Price & Ducker 1966: 261, figs 1–3. Skottsberg 1920: 277, figs 1–5. Womersley 1967: 236.
Thallus (sporophyte) (Fig. 42D) medium brown near apices (living) to dark brown (dried), very mucoid, usually 4–12 (–20) cm high with several axes from a small, discoid, rhizoidal holdfast 3–10 mm across; epilithic or on barnacles. Axes with several to numerous, radially and irregularly placed laterals, 2 mm-3 cm apart, 4–8 mm in diameter, basally constricted, becoming rugose (especially when dried), linear to clavate and usually increasing slightly in diameter to just below the rounded branch apex which has a slight depression. Clavate to pyriform "apical cells" (Fig. 44B), each tapering to a rhizoid, are present in the cortex of apical depressions (and young conceptacles) but these "apical cells" rarely divide (Clayton 1985). Growth (Fig. 44B) apical and peripheral, with anticlinal or oblique divisions in epidermal cells and periclinal divisions in subapical cells, haplostichous, developing a pseudoparenchymatous cortex with epidermal cells round to polygonal in surface view and 8–12 µm across, and a filamentous medulla of broader filaments (15–) 20–25 µm in diameter (forming a reticulum inside the cortex) and numerous hyphae 8–10 µ in diameter, becoming hollow centrally in each branch (Fig. 44A). Clavate paraphyses (Fig. 44B), basally meristematic, 6–12 cells long with dense physodes and few phaeoplasts, 8–10 µm in diameter above, occur near each apical depression but are lost from mature cortical cells; cortical cells with a single stellate phaeoplast [with a central or lateral (Asensi et al. 1977) pyrenoid] and numerous physodes.
Reproduction: Unilocular sporangia (Fig. 44C) ovoid to clavate, 100–160 µm long and 40–60 µm in diameter, borne in conceptacles scattered over the thallus, lying within the cortex and medulla and accompanied by uniseriate phaeophycean hairs; sporangia producing zoospores.
Type from South Africa (Koenig); in LINN, 1275.8.
Selected specimens: Point Sinclair, S. Aust., mid eulittoral (Womersley, 7.ii.1954; ADU, A19578). Pearson I., S. Aust., mid eulittoral (Specht, 17.ii.1960; ADU, A24514). Cape Willoughby, Kangaroo I., S. Aust., mid eulittoral (Womersley, 16.i.1946; ADU, A3161). Petrel Cove (Victor Harbor), S. Aust., mid eulittoral ( Womersley, 29.i.1950; ADU, Al2929). Point Bunbury, Apollo Bay, Vic., mid eulittoral (Womersley, 12.iv.1959; ADU, A22645). Cape Marengo, Vic., mid eulittoral ( Womersley, 13.x.1985; ADU, A56851). Koonya Bay, Mornington Pen., Vic., mid eulittoral ( Womersley, 15.i.1974; ADU, A44620-"Marine Algae of southern Australia" No. 146). Mallacoota Point, Vic., mid eulittoral ( Womersley, 12.xi.1982; ADU, A55341). Three Hummocks I., Tas., mid eulittoral (Wollaston & Mitchell, 21.ii.1964; ADU, A27631). Bicheno, Tas., mid eulittoral (Womersley, 19.i.1949; ADU, A10213). Goose I., Tas., lower (?) eulittoral (Bennett, 19.vi.1954; ADU, A19813). Probable microthallus: Sandy Cape, N.W. Tas., lower-mid eulittoral (Bennett, 15.i.1954; ADU, A19727).
Distribution: South Africa, Juan Fernandez I., New Zealand, and the more northern Sub-antarctic Islands.
In southern Australia, from Point Sinclair, S. Aust. to Newcastle, N.S.W. (May 1939, p. 206) and around Tasmania.
Taxonomic notes: Microthallus (gametophyte) (Fig. 44D, probable) densely tufted, 1–10 mm high, filamentous with basal cells and some lower cells of erect filaments developing longitudinal septa, cells with a single stellate phaeoplast and central pyrenoid (Fig. 44E), producing plurilocular organs (gametangia?) from which a further microthallial stage arises producing plurilocular sporangia and also macrothalli (sporophytes) (Price & Ducker 1966).
Splachnidium rugosum is present invariably in the mid eulittoral on moderate to rough-water coasts, usually only during summer months (Nov. to March) but on the Victorian and Tasmanian coasts plants may be found at other times of the year. The microthallus stage occurs in winter, as filamentous mats commonly among barnacles (Price & Ducker 1966, p. 268); that illustrated in Fig. 44D,E is regarded as "probable" since it agrees with the Price & Ducker description, the cells show typical stellate phaeoplasts with a central pyrenoid, and it occurred at the same intertidal level as Splachnidium.
The pyriform "apical cells" in the apical depressions and conceptacles still remain of unknown function. Skottsberg (1920) considered they never give rise to new cells and are distinct from the apical cells of adjacent filaments; he considered they are probably a parasitic species of Codiolum (Chlorophyta). Recently, however, Clayton (1985) has shown these cells (which are apparently cut off from surface cells) to have typical phaeophycean phaeoplasts and finds that they probably only divide when the thallus branches. They are clearly not normal apical cells which give rise to the thallus and should not be referred to as such; their function remains unknown.
The stellate phaeoplast of Splachnidium is similar to that of Scytothamnus (Delépine et al. 1976) and is distinct from that of all other Phaeophyta.
References:
ASENSI, A.O. & DELÉPINE, R. (1976). Ecophysiologie et cycle de développement de la Pheophycée australe Splachnidium rugosum (L.) Grev. Bull. Soc. Phycol. Fr. 21, 35–47.
ASENSI, A.O., DELÉPINE, R. & GUGLIELMI, G. (1977). Nouvelles observations sur l'ultrastructure du plastidome des Phéophycées. Bull. Soc. Phycol. Fr. 22, 192–205.
CLAYTON, M.N. (1985). A critical investigation of the vegetative anatomy, growth and taxonomic affinities of Adenocystis, Scytothamnus and Splachnidium (Phaeophyta). Br. phycol. 1 20, 285–296.
DELÉPINE, R., ASENSI, A. & GUGLIELMI, G. (1976). Nouveaux types d'ultrastructure plastidiale chez les Phéophycées. Phycologia 15, 425–434.
GREVILLE, R.K. (1830). Algae Britannicae. (Edinburgh.)
HARVEY, W.H. (1858). Phycologia Australica. Vol. I, Plates 1–60. (Reeve: London.)
KUCKUCK, P. (1929). Fragmente einer Monographie des Pheosporeen. Biol. Anst. Helgol. 17, 1–93.
MAY, V. (1939). A key to the marine algae of New South Wales. Part. II. Melanophyceae (Phaeophyceae). Proc. Linn. Soc. N.S.W. 64, 191–215.
MITCHELL, M.O. & WHITTING, F.G. (1892). On Splachnidium rugosum Grev., the type of a new order of algae. Phycol. Mem., Pt. 1, 1–10, Plates 1–3.
PRICE, I.R. & DUCKER, S.C. (1966). The life history of the brown alga Splachnidium rugosum. Phycologia 5, 261–273.
SKOTTSBERG, C. (1920). Remarks on Splachnidium rugosum (L.) Grev. Sven. bot. Tidskr. 14, 277–287.
WOMERSLEY, H.B.S. (1967). A critical survey of the marine algae of southern Australia. II. Phaeophyta. Aust. J. Bot. 15, 189–270.
The Marine Benthic Flora of Southern Australia Part II complete list of references.
Publication:
Womersley, H.B.S. (14 December, 1987)
The Marine Benthic Flora of Southern Australia
Part II
©Board of the Botanic Gardens and State Herbarium, Government of South Australia
Illustrations in Womersley Part II, 1997: FIGS 42D, 44.

Figure 42 enlarge
Fig. 42. A. Scytothamnus australis (ADU, A29994). B. Scytothamnus fasciculatus (ADU, A57210). C. Adenocystis utricularis (ADU, A55951). D. Splachnidium rugosum (ADU, A55341).
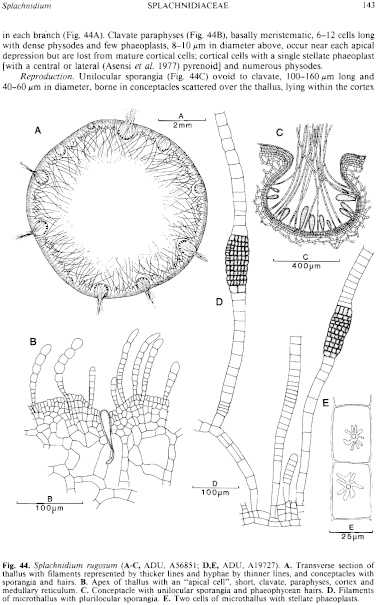
Figure 44 enlarge
Fig. 44. Splachnidium rugosum (A–C, ADU, A56851; D,E, ADU, A19727). A. Transverse section of thallus with filaments represented by thicker lines and hyphae by thinner lines, and conceptacles with sporangia and hairs. B. Apex of thallus with an "apical cell", short, clavate, paraphyses, cortex and medullary reticulum. C. Conceptacle with unilocular sporangia and phaeophycean hairs. D. Filaments of microthallus with plurilocular sporangia. E. Two cells of microthallus with stellate phaeoplasts.

|
Email Contact: State Herbarium of South Australia |

|









